《复杂生命的起源》梳理
Intro
这本书的结构实际上比较像综述,在谈到某一个问题的时候 Nick Lane 会先介绍一遍现有的理论,所以阅读过程中跟随他的思路去了解然后否定一个理论是比较有意思的。但是带来的问题也是阅读起来没有那么轻松,有时候会失去对主线的把握,所以在此对他提出的理论的主线进行时间上的梳理。
有机物合成的能量来源:碱性热液形成的质子梯度
根据 50 年代的「米勒-尤里实验」,我们已经知道无机物在闪电的能量输入下是可以合成出有机物的。但是在 Lane 看来,这里闪电提供的能量实在是太少了,要让一锅「原始汤」里面有机物的浓度足够高,靠闪电是不够的,他更倾向于能量的来源是碱性热液喷口形成的质子梯度。
当地下水渗透到地壳深处的时候,水会和橄榄石(一种含有镁和铁的硅酸盐)反应,主要产生蛇纹石和氢气,这个过程中也会生成氢氧化镁。所以这个整个过程就会生成富含氢气的碱性热液。
我们知道在一般的情况下,氢气和二氧化碳是不能直接反应的,因为在 pH 相同的情况下,氢离子的还原电位是小于甲酸根的,所以会更容易得到电子,于是氢原子就不会失去电子。而增加 pH 只会同步增加二者的还原电位,让反应一直不会发生,所以说这个反应虽然是一个放热的反应,但是存在着一个起始的壁垒。但是碱性热液环境营造了一个非常不一样的场景:硫化亚铁这样的无机催化剂形成的无机物薄壁可以让两端有不同的质子浓度,从而使得两边的还原电位关系倒转,驱动这个反应自发进行。于是,在碱性热液中,就可以存在持续的生成有机分子,比如甲烷、甲醇等。
LUCA 的出现
在碱性热液和温度更低的海水融合的过程中,会有「热泳」(thermophoresis) 的现象,使得有机物分子(尤其是其中的大分子)被聚集在一个比较小的范围中,形成一种浓缩的效果。在这样的环境下,氨基酸可以聚合成蛋白质,脂肪酸可以聚集成一个个囊泡,而核苷酸也可以合成 RNA。就是在这里,产生了著名的 LUCA (Last Universal Common Ancestor),也即作为定义的生命的共同祖先。值得注意的是 LUCA 不一定是第一个生命,而只是最后一个共同的祖先,这二者未必是同一的。
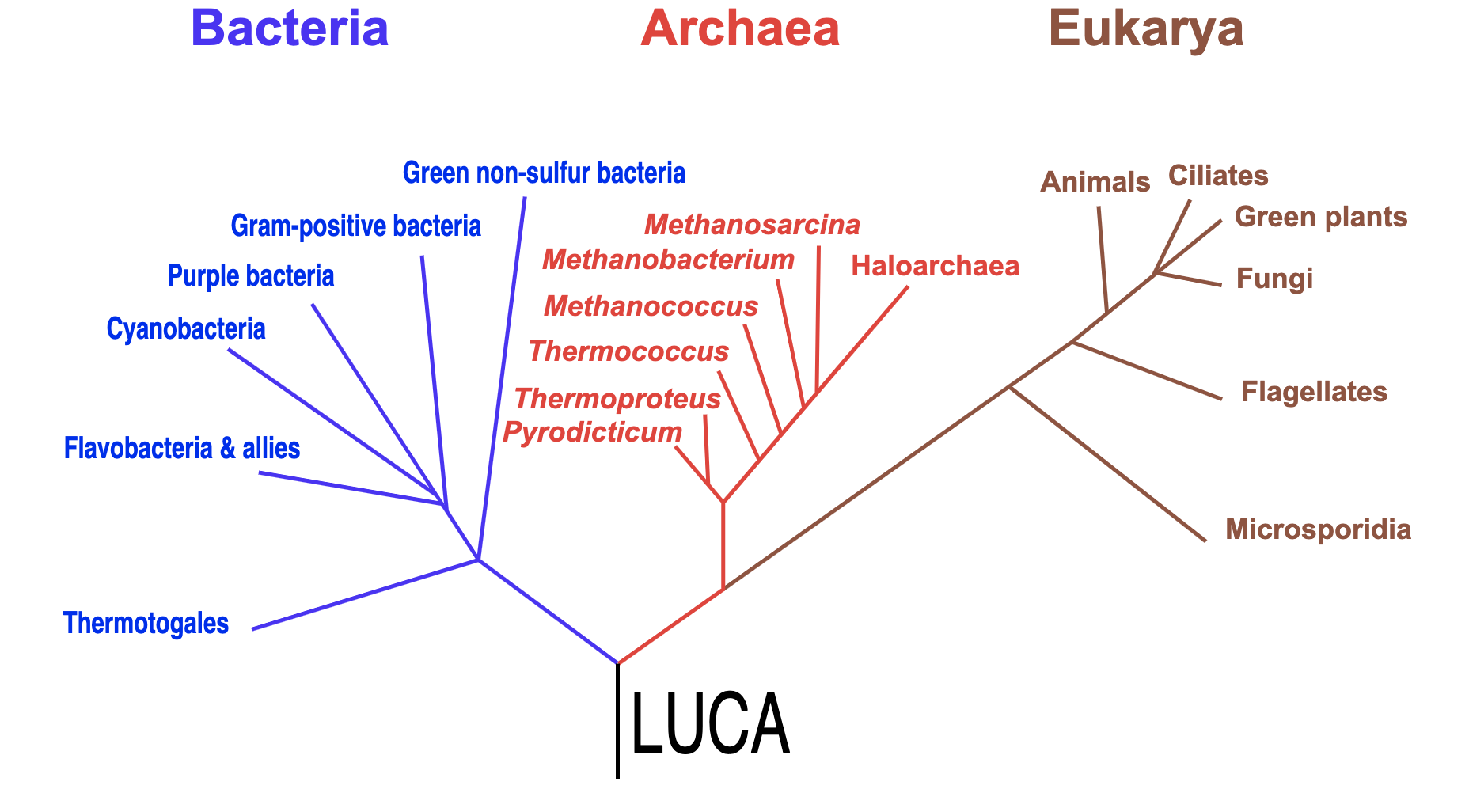
这时候的 LUCA 是怎么样的呢?实际上它显然仍然只是一团有机物的聚合,同时可以利用质子梯度来合成有机物。只是符合了我们对活的细胞的 6 个特征的描述:能量流、碳元素流、有用以催化的无机物、有遗传物质复制的能力、有区隔化(囊泡)、排泄。可以想象它大概是一个卡在无机薄壁上的囊泡,里面主要装了一些蛋白质,而这个囊泡(细胞膜)是比较稀疏的,可以让质子流轻松流入和流出。在这个流入和流出的过程中,可以通过细胞膜上的 ATP 合酶来合成 ATP。然后 LUCA 再利用 ATP 来合成更多的蛋白质和遗传物质,以达成自我复制。
离开喷口:反向转运蛋白的出现
到这个时候,LUCA 的细胞膜一定是要可以让质子渗漏的,因为如果进入细胞内的氢离子没能即时离开细胞内,就会让梯度消失。也就是说在这里质子的流向应该是从外部环境通过细胞膜(ATP 合酶)进入细胞内,然后再透过细胞膜扩散到外部环境中。这里质子进入细胞的动力是电化学梯度,而扩散出细胞的动力则是通过环境的对流,让质子自然渗透出去。所以在这种环境下,实际上细胞是没有任何的自然选择动力去降低细胞膜的渗透率的。除非它有了某种机制,可以主动泵出离子。
这个时候钠-质子反向转运蛋白 (Na⁺/H⁺ antiporter) 的出现完成了这一历史使命,其作用就是耦合离子梯度的能量转换,通过泵入一个钠离子来泵出一个质子,或者是反向,通过泵入一个质子来泵出一个钠离子。这种机制有重大的意义:
- 反向转运蛋白可以利用质子梯度来驱动泵出钠离子。因为质子是远比钠离子更容易通过细胞膜的,所以泵入的质子可以渗漏出去,然后持续来驱动泵出,而泵出的钠离子就会持续停留在细胞膜之外,维持钠离子的梯度。也就是说把质子梯度转化成了更加稳定的钠离子梯度
- 同时,这种转化并不是就消灭了原有的质子梯度,质子梯度仍然能工作。所以有反向转运蛋白相当于是给细胞提供了额外的能量,书里面说计算是多了 60% 的能量。这里额外的能量可以让细胞在 pH 差更小的环境中生存,原先可能是 pH 差 3,现在可以在 pH 相差 2 以内的环境生存
所以至此,细胞已经可以在更加广大的范围中生存,比如扩散到周边的喷口。但是显然,因为能量的最终来源仍然是质子梯度,所以细胞仍然不能离开喷口。但是反向转运蛋白的出现让细胞有了新的选择压力
- 降低细胞膜的渗透率,让钠离子可以更多的留在细胞外,这样就能提高能量的利用率
- 同时,主动去泵出质子也有了意义,之前的时候泵出就会直接渗漏回来,而现在可能会通过反向转运蛋白回来,这样就能多泵出一个钠离子。虽然这不是一定的,但是主动泵出质子就有可能带来更多能量,所以会驱动质子泵的演化。而有了质子泵之后,有更加能驱动降低细胞膜的渗透率,因为可以让更多的质子通过反向转运蛋白流回来,而不是渗漏回来
到这里,有了质子泵和更加不渗漏的细胞膜,细胞膜就有了可以离开喷口的能力。本质上来说,就是把对于外部的质子梯度的依赖转化成了一种自己能够制造的梯度的依赖,有了一种能量上的自主权。
内共生:复杂生命的出现
首先可以思考,为什么细菌和古菌没能把细胞做大——原因在于能量密度。单个原核细胞只有一层质子驱动的呼吸膜,体积一旦增大,表面积∶体积比急剧下降,能量供应跟不上基因复制和蛋白合成的开销。Nick Lane 把这称作「每个基因可支配的能量」(energy per gene)的硬限制。
大约 20 – 18 亿年前,一支与现代古菌相似、已能进行吞噬(原始吞噬作用)的宿主细胞,误把一只 α-变形细菌留在了体内。它没被消化,而是继续用自己的内膜做呼吸,形成了原始线粒体。此举把呼吸膜从「单层包壳」外移到「无数颗内置反应堆」,把能量生产从面积限制变成可线性扩充的并联结构。一旦细胞需要更多能量,只要让线粒体分裂即可,宿主基因组却不必同步增大——这正是突破能量密度瓶颈的关键。
线粒体规模膨胀,但它们的基因却在随后几亿年里剥离、转移到真核核基因组,只保留少量核心电子传递蛋白的编码基因,以便在原位快速调节呼吸链。结果是真核细胞获得了一个「低成本、可无限扩容」的发电外包系统:
- 线粒体 DNA ≈ 1% 甚至更少,而呼吸膜面积却可增长几个数量级
- 宿主核基因组因此可以负担更多调控元件、剪接体、信号通路,而不用担心能量赤字。Lane 估算,真核细胞 energy per gene 比典型细菌高 1,000 – 10,000 倍,这才支撑得起长链的内含子、复杂的蛋白折叠监控和庞大的细胞骨架网络。
性的作用
这里还有一个非常有意思的点是内共生与「性」的出现可能高度相关。内共生的状态要求线粒体和整个细胞紧密合作,能量供给的量要匹配整体。但是因为线粒体有自己的复制节奏,突变速率比核基因高(应该和高强度进行氧化还原反应有关),所以如果是无性繁殖的话核基因会跟不上线粒体基因的突变节奏,久而久之就会出现失调的情况。
而性的出现解决了这个问题:生殖过程中只使用一方的线粒体基因(通常是雌性的),这样就固定了要匹配的「靶」,然后重组双方的核基因去匹配这个靶。这样就更容易找到和线粒体基因所匹配的新的核基因(那些不适合的有自然选择帮忙去淘汰掉)。
LECA: 最后真核共同祖先
约在 16 – 15 亿年前出现的 LECA(Last Eukaryotic Common Ancestor)已经拥有今天绝大多数真核共有的装备:
- 成熟的线粒体 + ADP/ATP 转运蛋白
- 带核膜的基因组、真核式启动子与剪接体
- 动力蛋白、微管和肌动蛋白、微丝构成的可重塑骨架,支持酶运输和细胞分裂
- 高保真有丝分裂装置、减数分裂和两性循环
- 内膜系统(高尔基体、内质网、溶酶体)与吞噬-分泌通道
换言之,真核复杂细胞的整体模板在一次历史事件后几乎一蹴而就,而后 15 亿年的演化更多是在这套模板上做局部改装。
复杂生命是偶然的还是必然的
应该说二者都有,偶然性主要在于细菌进入古菌的成功内共生仅见一次;无第二个事件,所以也无第二套复杂生命。
必然性在于,只要能突破能量瓶颈,后续一系列结构与遗传创新几乎是自然选择的必然。
Lane 因此推测,宇宙里「简单生命」或许稀松平常,而「复杂生命」则需要极罕见的热化学、膜生化与内共生三重偶然叠加。